


“If these creatures did not exist, they would be almost too incredible to believe »
“Si ces créatures n’existaient pas, elles seraient trop difficiles à imaginer tant elles sont incroyables“
Sean B. Carroll

L’Ordre des Odonates est l’un des plus vieux ordres d’insectes actuels. Il fait partie du super ordre des Odonatoptères apparu au Carbonifère (300 Ma) avec l’ordre des Méganisoptères, insectes volants les plus grands ayant vécu sur Terre (plus de 70 cm de largeur pour Meganeura monyi). Les Odonates primitifs avaient déjà des caractères morphologiques proches des Odonates actuels : des yeux spécialisés, quatre grandes ailes et une spécialisation des pattes pour la capture de proies en vol. Ceci suggère un comportement de prédation aérienne des premiers Odonates semblable à celui des Odonates actuels (Nel et al. 2018).
Malgré l’ancienneté de l’ordre, les Odonates présentent une remarquable stabilité sur le plan anatomique. Cette caractéristique peut suggérer qu’ils ont rapidement atteint une forme optimale de spécialisation pour la prédation aérienne garantissant une efficacité d’adaptation durable.
Des observations spectaculaires ?
Les libellules sont de grandes prédatrices, agiles et rapides, inféodées aux zones humides. Chez les anisoptères (libellules vraies) les deux principales familles possèdent des différences comportementales à l’état imaginal. Comportementalement parlant, les Libellulidés ont un type de chasse actif “Sit and wait” : elles se posent sur une tige, une branche ou tout support pouvant les placer au-dessus de l’eau. Elles attendent de détecter une proie pour ensuite la pourchasser en volant et revenir à leur emplacement de départ pour la manger. Ces séquences se font souvent en moins de 0,5 s à 1 s. Quant aux Aeschnidés, ils possèdent un type de comportement de chasse de type “Hawker” : ils « patrouillent » en vol, chassant, attrapant et mangeant les proies qui passent à proximité.
Le premier type de comportement est facile à observer quand on s’arrête près d’une mare ou d’un étang. Le second demande un suivi sur une large zone et n’a pas de séquences bien visibles ; il faut avoir l’œil averti pour le distinguer. De fait, les études comportementales sont principalement faites sur des libellules de la famille des Libellulidés.
Dans les années 2000, plusieurs études sur l’observation comportementale de chasse des libellules paraissent. Elles ont tenté de déterminer le taux de succès de chasse des libellules, plus précisément de celles du genre Libellula appartenant à la famille des Libellulidés. Mais qu’est-ce qu’un « taux de succès » ? Un taux de succès se calcule en divisant le nombre de tentatives réussies de capture d’une proie par le nombre de tentatives total. Si un prédateur arrive à attraper 15 proies sur 20 tentatives alors il a un taux de succès de chasse de 75%.

Pour étudier ce comportement, il existe plusieurs méthodes qui ont évolué au fil des avancées technologiques. La première d’entre elles, la moins coûteuse et la plus ancienne, consiste en l’observation du comportement dans le milieu naturel. Elle se fait alors directement à l’œil nu ou à l’aide de jumelles. Les premières études connues ont été faites dans un milieu naturel, rendant difficile l’acquisition des données. Le taux de succès s’élevait alors à 76% (Baird and May, 1997). Avec les avancées technologiques, l’utilisation de la capture vidéo ralentie couplée à la création de milieux naturels artificiels en laboratoire a permis de faciliter les observations. C’est grâce à ces techniques que peu après 2000, des chercheurs ont réussi à observer un taux de succès de 97% (Olberg et al, 2000) ! C’est un taux de succès considérable, surpassant la majorité de ceux des prédateurs mammifères ou aviaires. Par exemple, le guépard est à 58,2% et le faucon pèlerin, qui évolue dans un environnement similaire en trois dimensions, atteint seulement 34,9% (Roalkvam, 1985).
Pourquoi un taux de capture aussi élevé ?
Cette efficacité peut s’expliquer par les besoins physiologiques des libellules : à l’état adulte elles doivent gérer un équilibre coût/bénéfice très précaire car leur besoin énergétique est énorme. Pour chaque tentative de capture d’une proie, la libellule consomme énormément d’énergie et se met en danger par rapport à ses prédateurs. Elles doivent donc méticuleusement choisir leurs proies et leurs attaques.
Chez les libellules, le principal besoin énergétique est dédié au vol. Chez le Libellulidés Plathemis lydia, il a été relevé que plus de 60% de la masse musculaire des mâles est dédiée aux muscles responsables du vol (Marden 1989). Une différence sexuelle existe : les mâles ont un besoin très élevé en nourriture car ils passent la majeure partie de leur temps en vol pour défendre leur territoire face à leurs concurrents ou pour s’accoupler. Les femelles, quant à elles, ont des besoins énergétiques principalement dédiés à leurs organes reproducteurs.
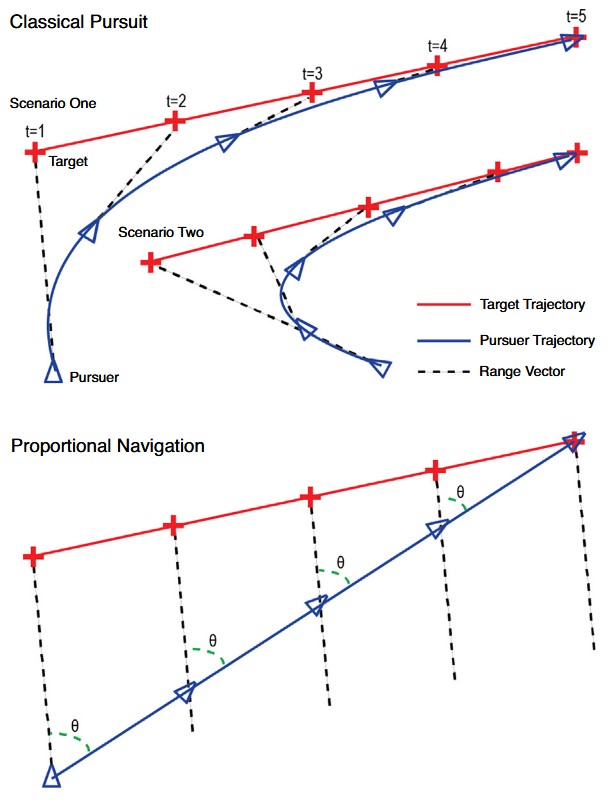
Des maîtres de l’interception
Pour comprendre comment les libellules réussissent à atteindre un taux de capture si élevé, il faut analyser leur manière d’attraper leurs proies. Dans le règne animal, les prédateurs actifs peuvent attraper de plusieurs façons une proie ou un objet qui se déplace. La première est la poursuite, à la manière d’un chien et d’une balle ou d’un guépard et d’une gazelle : le prédateur va dans la direction directe de la proie avec pour but de diminuer la distance entre lui et cette dernière pour ensuite asséner un coup fatal. Très souvent cette technique produit une trajectoire en forme de spirale.
La seconde manière utilisée par des prédateurs actifs, plus complexe et performante, est la technique d’interception de la proie. C’est cette technique que les Libellules utilisent. Elle n’a pas pour but de diminuer la distance entre la proie et le prédateur mais d’anticiper un point dans l’espace ou les trajectoires de la proie et du prédateur vont se rencontrer. Les libellules anticipent le point de rencontre de leur trajectoire avec celle de leur proie en allant non pas dans la direction de la proie mais dans la direction de la trajectoire de la proie. Ce comportement implique une capacité de prédiction de la position future de la proie, ce qui suggère une complexité comportementale qui n’est pas commune chez les insectes compte-tenu de leur faible nombre de neurones (Olberg et al., 2000).

.){kind=link}
D’immenses yeux de chasseuse
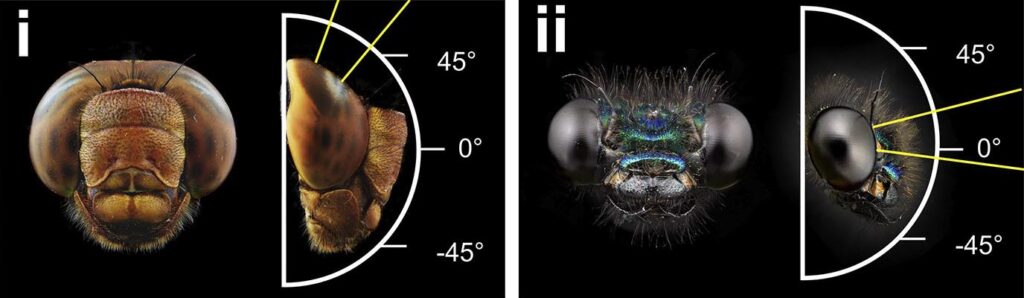
Si on remarque bien quelque chose chez les libellules, ce sont leurs grands yeux complexes occupant presque toute leur tête. Les libellules possèdent en effet les plus grands yeux de la classe des insectes en nombre de facettes/ommatidies : on en a comptabilisé jusqu’à 30 000 (29 247) chez des Aeschnidés contre 1500 chez les drosophiles par exemple. Pourvues ainsi d’une vision à 360° cumulée à une articulation particulièrement souple entre la tête et le thorax, les libellules peuvent avoir une appréhension parfaite de leur environnement. Les chercheurs ont aussi montré que les libellules possédaient des yeux organisés en régions spécialisées, liées à leurs comportements. Il existe ainsi une zone d’acuité visuelle maximale appelée fovéa, qui se situe à environ 55° au-dessus de l’horizontal et qui forme un croissant partant de la ligne médiane – qui est la zone où s’unifient les deux yeux composés – et allant jusqu’à la périphérie. Cette zone dorso-frontale peut être particulièrement visible chez certaines espèces de libellules par le changement de couleur des ommatidies/facettes. Cette couleur différente est due à une richesse particulière en pigments sensibles aux longueurs d’ondes courtes (540nm-bleu), ce qui permettrait de reconnaître les proies plus facilement par l’effet d’un contraste entre le fond bleu du ciel et la proie. Cette région confère donc un avantage important pour le comportement de chasse car elle permettrait aux libellules de particulièrement bien voir leur proie.
Chez l’humain cette zone existe aussi et est utilisée en permanence. Elle correspond à la zone qui nous permet de voir l’image la plus nette, mais étant donné les particularités physiologiques et comportementales des libellules, elles ne l’utilisent que lorsqu’elles chassent !
Des « moteurs » très performants
Les libellules (Odonates), font partie des Paléoptères qui sont caractérisés par deux paires d’ailes indépendantes. Les quatre ailes sont membraneuses, fines, longues, larges et traversées par un fin réseau de veines ou nervures tubulaires laissant passer l’hémolymphe, des systèmes trachéens et des neurones. La morphologie des ailes des libellules a très peu changé au fil des 300 derniers millions d’années, signe que leur morphologie alaire est bien adaptée à leur comportement ainsi qu’à leur habitat.
Elles possèdent deux paires de muscles dorsaux-ventraux insérés directement sur les plaques axillaires et humérales. Les muscles abaisseurs et les muscles élévateurs provoquent un mouvement de rame de haut en bas, générant de la portance. Leur spécificité anatomique d’avoir les muscles directement insérés sur les ailes permet de battre des ailes indépendamment l’une par rapport à l’autre et confère aux libellules les dynamiques de vol les plus diversifiées et les plus précises de tous les insectes.
Capables de battre des ailes à des fréquences et à des angles différents, les libellules peuvent voler en avant, en arrière (spécificité rare dans le règne animal), vers la gauche ou la droite et vers le haut ou le bas. Grâce aux mouvements asynchrones de leurs ailes, les Anisoptères peuvent changer de direction rapidement et même planer. Les libellules possèdent l’une des masses musculaires dédiées au vol les plus importantes du règne animal, certaines espèces – comme la libellule globe-trotteur (Pantala flavescens…- pouvant même migrer d’un continent à l’autre par-delà des mers, des montagnes et des déserts.
Des neurones bien câblés
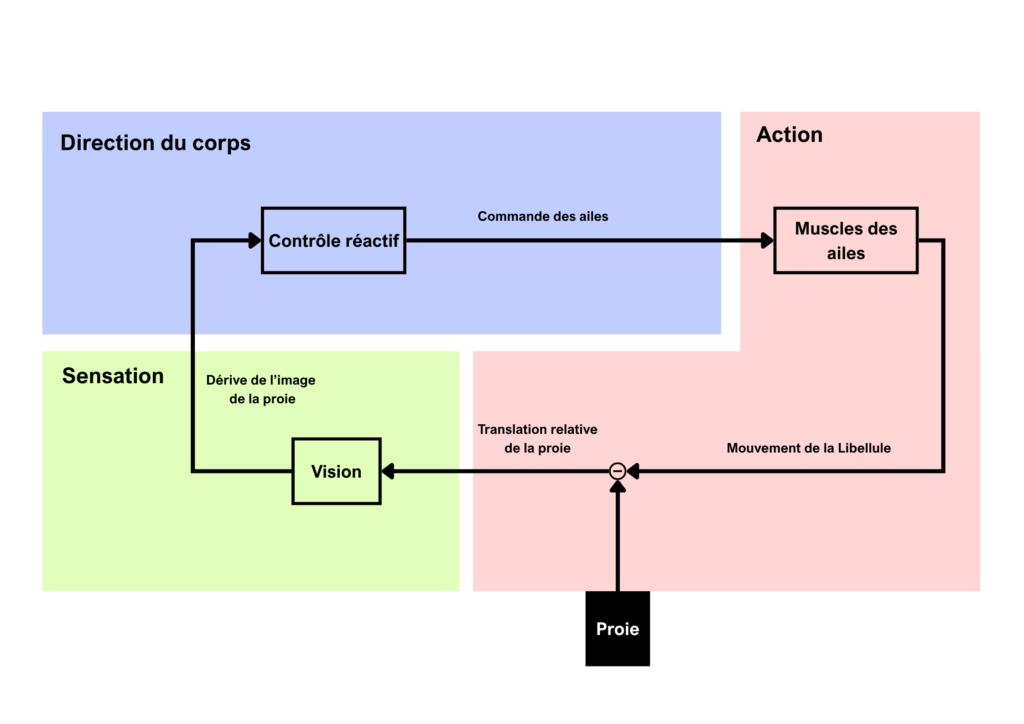
Pendant les observations de comportement de chasse, les chercheurs ont découvert que les libellules mettaient seulement 30 à 50 millisecondes à corriger leurs trajectoires d’interception lorsque la proie changeait brusquement de direction… ce qui a été une première piste pour expliquer un taux de succès de chasse si élevé ! Mais la suite de l’étude a révélé d’autres surprises !
Vingt ans plus tôt on avait découvert un certain type de neurones nommés TSDN pour “Target Selective Descending Neurons “ ou Neurone Descendant à Cible Sélective. Au nombre de huit paires, ce sont des interneurones qui ont des fibres descendantes. Ils sont situés au niveau du protocerebrum passant par la corde nerveuse ventrale et irriguant les ganglions thoraciques (prothoracique, mésothoracique, métathoracique) responsables de l’activité musculaire provoquant les battements d’ailes. Lorsqu’on les stimule électriquement de manière artificielle, cela provoque des micros-changements de positionnement d’ailes ce qui a été interprété comme des changements de direction. Lorsque les chercheurs recherchaient alors des voies neuronales à faible latence pouvant expliquer ce comportement, les neurones TSDN étaient alors de bons candidats.
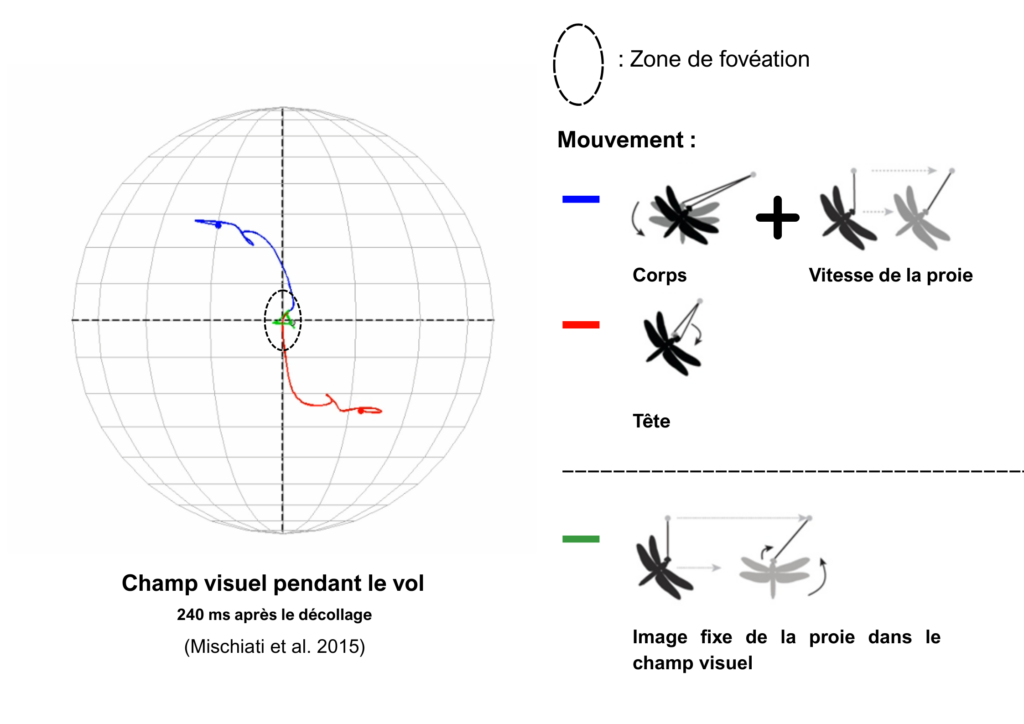
Avec les avancées technologiques de suivi visuel informatique, on a pu tester par la suite la réactivité de ces neurones non plus avec des stimuli électriques mais avec des proies artificielles composées de perles attachées à une longue tige en métal pouvant déclencher une séquence de chasse. Avec cette méthode, les chercheurs confirmèrent la faible latence entre le changement de comportement de vol de la proie et la réactivité de la libellule, environ 29 ± 6,4 millisecondes en moyenne, et chose nouvelle, on observa que la tête des libellules suivait en permanence la proie pendant le vol. La libellule faisant en sorte d’orienter sa « mire », zone formée par la ligne médiane de ses deux yeux et la zone de haute acuité visuelle située entre 50 à 60 ° au-dessus de l’horizon directement vers la proie (Olberg et al. 2007).
Les chercheurs ont encore creusé davantage la voie des interneurones TSDN et ont alors trouvé qu’ils s’activaient uniquement lorsqu’un stimulus visuel était détecté dans la zone de forte acuité. Et plus intéressant encore, les TSDNs s’activaient seulement lorsque la proie sortait d’une certaine zone de la forte acuité, appelée zone de fovéation, ce qui signifiait que la proie avait changé de direction (Gonzalez-Bellido et al. 2013).
Ainsi, tant que la proie est fixe au niveau de la zone de fovéation dans le champ de vision de la libellule, la libellule vole à la rencontre de la trajectoire de la proie et va donc pouvoir l’attraper. Mais si la proie n’est plus fixe dans son champ de vision et sort de la zone de fovéation, alors les interneurones TSDNs sont capables de détecter cela et de corriger la trajectoire de la libellule en stimulant les motoneurones musculaires des ailes des ganglions thoraciques pour lui permettre de se replacer dans une trajectoire d’interception. Les chercheurs décrivent donc le comportement de chasse des libellules comme un comportement de réactions aux mouvements de la proie.
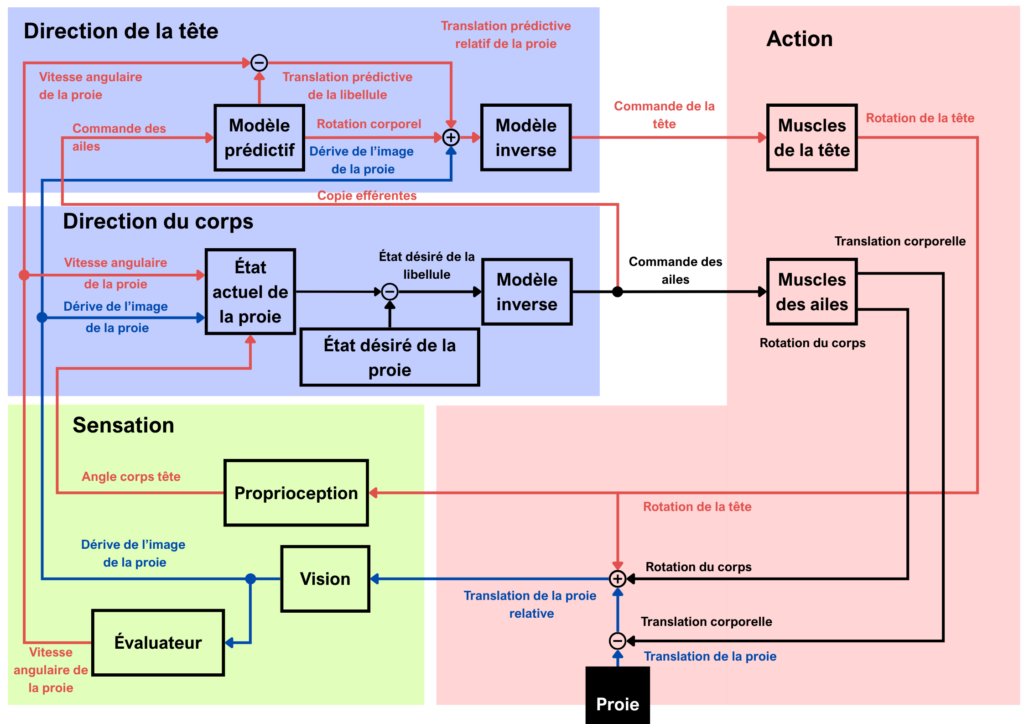
Les mouvements de la libellule et de la proie modifient la vitesse angulaire et la position de la proie, ce qui entraîne le prochain cycle de pilotage. Modèle conceptuel résumant une proposition de l’architecture de contrôle utilisée par la libellule, une voie de pilotage prédictive (rouge) et une voie de pilotage réactive pilotée visuellement (bleu) (Mischiati et al., 2015)
Cependant, l’étude de Mischiati et al. parue en 2015 dans Nature remet en cause vingt années de modèle réactif. L’utilisation d’enregistrements vidéo ainsi qu’un marquage rétro-réfléchissant sur les libellules a permis d’observer les différents angles de la tête et du thorax pendant le décollage et le vol d’interception. Avec des proies artificielles gérées par ordinateur, augmentant donc les possibilités de l’étude de réactions aux variations de vitesse. Leurs résultats montrent que lorsque les libellules décollent, elles placent leur corps dans la direction de la trajectoire de la proie. Ce comportement permet de supprimer la dimension gauche-droite de la trajectoire d’interception car il ne lui resterait plus que les dimensions haut-bas et avant-arrière à gérer, ce qui facilite l’interception.
Un problème apparaît cependant lorsque la libellule pivote son corps dans le plan horizontal mais que sa tête garde la proie en ligne de mire, car cela entraîne une aberration visuelle du mouvement de la proie qui pourrait mener à une mauvaise estimation de la trajectoire de la proie. Pour contrer cela, la libellule annule l’aberration grâce à des mouvements contraires de la tête et cela se fait en moyenne en 4 ± 4 ms.
Il faut savoir que chez les insectes, la seule activation des muscles prend en moyenne 5 ms et la réaction la plus rapide à un stimulus met environ 30 ms à activer une réaction musculaire comme nous l’avons vu plus tôt. Donc les mouvements de têtes sont bien trop rapides pour que cela soit un comportement réactif. La tête produit ainsi des mouvements prédictifs pour annuler les aberrations visuelles créées par la rotation du corps et le mouvement de la proie. Les chercheurs concluent que les libellules disposent de modèles internes prédictifs pour annuler les mouvements pouvant induire des changements visuels compromettant la fovéation (mécanisme permettant de garder la proie dans la zone de fovéation) et donc l’interception de la trajectoire de la proie. De plus, un autre indice d’un modèle interne prédictif est que lorsque les proies artificielles augmentent subitement leur vitesse de 80 % pendant le vol d’interception, cette augmentation n’a pas d’effet visuel discernable sur la trajectoire de la libellule. Ils en concluent que les libellules ne réagissent pas à l’augmentation brutale de vitesse de la proie mais intègre cela dans leurs modèles internes prédictifs.
Cependant, les TSDNs ne sont pas inutiles car une voie réactive existe toujours. En particulier, ils interviennent lors des mouvements brusques et des manœuvres d’évasion de la proie (Mischiati et al. 2015).
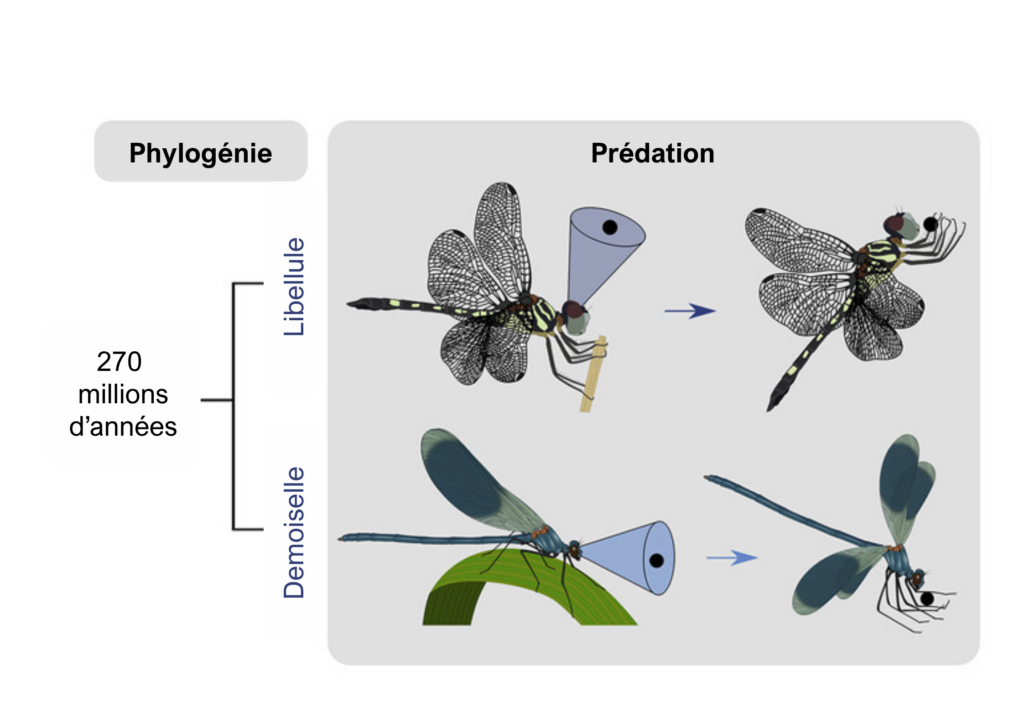
Et chez les demoiselles ?
Les demoiselles (Zygoptères) ont un ancêtre commun avec les Anisoptères il y a environ 270 millions d’années, cependant elles restent morphologiquement proches avec quelques spécificités, comme, outre des différences larvaires, la fermeture des ailes à la verticale du thorax lorsque qu’elles sont au repos, des yeux non holoptiques et des mécaniques de vols beaucoup moins vifs et agiles. Avec les mêmes méthodes, les chercheurs ont essayé de retrouver si ces neurones sont aussi présents chez les demoiselles, et ils ont réussi à démontrer que ces dernières possédaient aussi des interneurones TSDN. La différence entre les deux sous-ordres est le comportement de la tête pendant le vol. Les Anisoptères fixent leurs proies avec leur « mire » située dans leurs zones de forte acuité (50°/60°) alors que les zygoptères fixent les proies avec un angle allant de 3,9° à 11,4° par rapport à l’horizon, ce qui est compatible avec la chasse des demoiselles qui est de type frontal. Autre différence, pour activer les TSDN, les demoiselles ont besoin de recevoir des stimulations provenant des deux hémisphères oculaires, alors que chez les libellules, une stimulation d’un hémisphère oculaire suffit à activer les TSDN de la zone. Malgré ces différences, anatomiquement et physiologiquement, les TSDNs des deux sous-ordres sont très similaires. Les chercheurs suggèrent que ce caractère homologue devait faire partie d’un caractère partagé avec leur ancêtre le plus proche, il y a environ 270 millions d’années (Supple et al. 2020).
Ce caractère ancestral pourrait être recherché dans le genre Epiophlebia, originaire du sous-continent indien, d’Asie de l’Est et d’Asie du Sud-Est et curiosité parmi les Odonates. En effet, c’est le seul genre vivant de « libellules-demoiselles » à n’appartenir ni à la famille des libellules ni à celle des demoiselles tout en en présentant des caractères intermédiaires.
Pour en savoir plus
Marden J. H., 1989 – Bodybuilding Dragonflies: Costs and Benefits of Maximizing Flight Muscle, Physiological Zoology 62, no 2https://www.journals.uchicago.edu/doi/10.1086/physzool.62.2.30156182
Nel A. et al., 2018 – Palaeozoic giant dragonflies were hawker predators. Sci Rep 8, 12141
Roalkvam R. 1985 – How effective are hunting peregrines ? Raptor Research, 19(1), pp.27–29