Sauf mention contraire, les illustrations sont de l’autrice Isabelle Avisse.
Programme de sciences participatives Spipoll
L’analyse de photographies d’insectes collectées lors de sessions de sciences participatives révèle parfois des curiosités dignes de véritables enquêtes. Voici comment une collection de photos prises au fond du jardin a permis de révéler des liens fonctionnels entre certains insectes et certaines plantes.
Entomologiste amatrice, je contribue tous les ans au programme de sciences participatives Spipoll, acronyme de « Suivi photographique des insectes pollinisateurs ». Chapeauté par l’Opie (Office pour les insectes et leur environnement) et le MNHN (Muséum national d’histoire naturelle), le protocole de suivi dudit programme de sciences citoyennes incite ses contributeur·ice·s à photographier durant vingt minutes tous les insectes vus sur une même espèce végétale en fleur avant de télécharger les photos obtenues sur le site Internet dédié et d’identifier les insectes immortalisés à l’aide des outils proposés, dont une clé en ligne, permettant de distinguer 630 espèces ou groupes d’espèces indiscernables sur photo. D’amplitude nationale, ce suivi photographique vise à obtenir des données quantitatives sur les pollinisateurs et autres insectes floricoles en France afin de mesurer les variations de leur diversité, mais aussi de la structure des réseaux de pollinisation sur l’ensemble du territoire. Les collections photographiques ainsi créées alimentent une base de données précieuses pour nombre de chercheurs et de scientifiques.
Mes observations de juin 2023
Mon jardin (cliché ci-dessus) se situe à une trentaine de kilomètres au sud de Caen, entre les villes de Thury-Harcourt et de Falaise (Calvados). À la mi-juin 2023, alors que j’y photographiai – dans l’objectif de télécharger une collection « Rue officinale » (Ruta graveolens, Rutaceae) sur le site Internet du Spipoll – les insectes qui visitaient un pied de cette belle plante aromatique fleurant l’orange et la noix de coco, je capturai en images deux abeilles sauvages femelles nommées andrènes (genre Andrena, famille des Andrenidae) porteuses de pollinies d’orchidées aux couleurs différentes fixées au front.
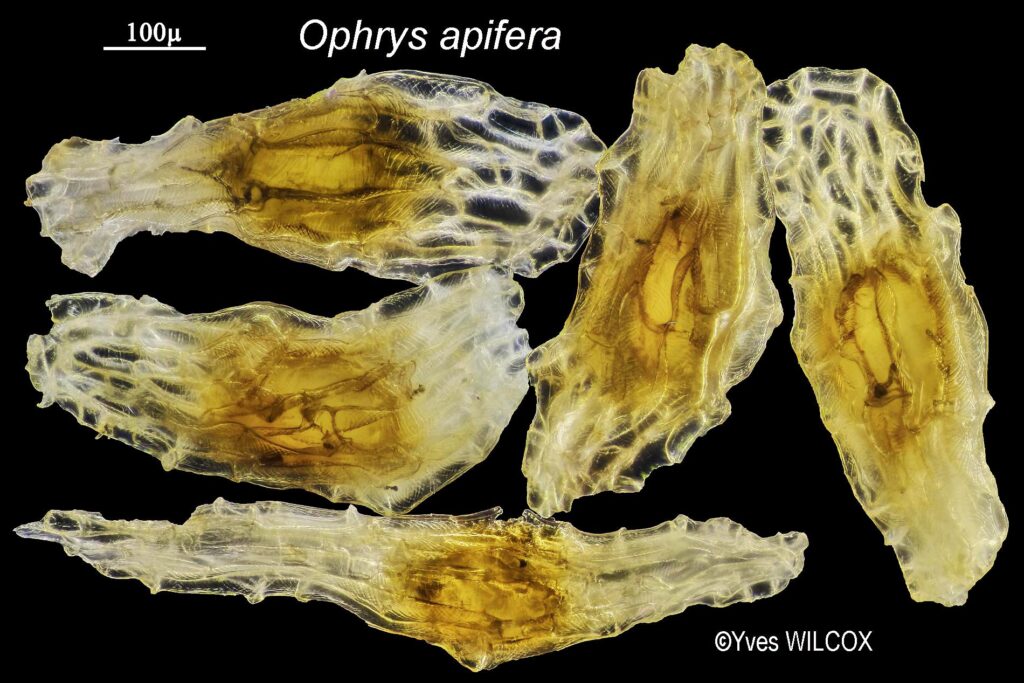
Sur ces clichés présentant les deux Andrènes, les massulae1 des pollinies (masses cohérentes de grains de pollen) sont abîmées, ce qui indique que les andrènes ont visité plusieurs autres fleurs et y ont déposé du pollen.
À la même période, sur une inflorescence de lavande (Lavandula sp., Lamiaceae) plantée à une dizaine de mètres dudit pied de rue officinale, je photographiai une autre femelle d’andrène – semble-t-il de la même espèce (Andrena flavipes, l’andrène à pattes jaunes, abeille polylectique commune dans les jardins et paysages agricoles) – également chargée de pollinies collées au même endroit du corps, des pollinies distinctes par la couleur de celles précédemment photographiées.
À quelles espèces d’orchidées pourraient bien appartenir ces 3 morphotypes de pollinies ?…
Avant de poursuivre et de développer cette interrogation, quelques mots tout de même sur les andrènes, véhicules des organes reproducteurs mâles des 3 types d’orchidées mystères photographiés dans le contexte précédemment décrit.
Les andrènes
Les Andrénidés sont une famille d’abeilles solitaires largement répandue à la surface du globe, dont les États-Unis d’Amérique possèdent le plus grand nombre d’espèces (un peu plus de 1 200 décrites). En revanche, ni l’Australie ni les régions tropicales de l’Asie n’en ont de représentantes. Les andrènes sont majoritairement présentes dans les aires tempérées et arides du globe. Dites « minières », « fouisseuses » ou « des sables », ces hyménoptères Apoïdes anthophiles (Anthophila, anciennement « apiformes ») regroupent des espèces qui ne nidifient que dans le sol. Elles sont soit polylectiques soit oligolectiques : généralistes, les abeilles polylectiques butinent une grande variété de fleurs ; spécialisées, les abeilles oligolectiques ne récoltent de pollen pour alimenter leur descendance que sur quelques espèces de plantes d’un même genre ou d’une même famille botanique. Au moins182 espèces d’andrènes ont été recensées en France (Ropars et al. 2025).
Divers morphotypes d’andrènes femelles qui fréquentent jardins et milieux agricoles dans le Calvados
À l’exception de quelques espèces d’Andrena typiques, l’identification spécifique des abeilles de ce genre ne saurait s’effectuer sur la base de photographies, car nombre d’espèces sont morphologiquement très ressemblantes les unes des autres. Les tuer avant de les préparer pour les analyser sous la loupe binoculaire à l’aide de clés de détermination idoines est aujourd’hui encore indispensable au travail des taxonomistes (dont je ne suis pas).
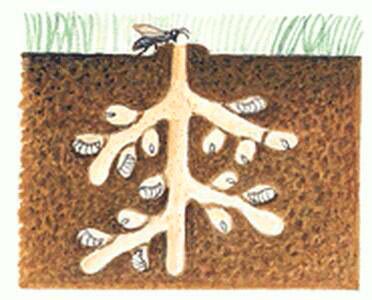
Terricoles, les femelles d’andrène aménagent des terriers qu’elles creusent dans le sol au sein de zones à végétation rase ou clairsemée, de prairies permanentes, de lits de route sèche, de chemins sablonneux… La femelle fore un long tunnel à ramifications multiples s’achevant en chambres larvaires (une dizaine environ par terrier) ; dans chacune d’elles, elle stocke une boulette de pollen mêlé de nectar (pain d’abeille). Après avoir pondu un œuf sur chacune de ces boules, elle scelle chacune des cellules à couvain. Les larves consommeront cette nourriture jusqu’à la nymphose.
Bien que les nids soient parfois construits à proximité les uns des autres (en « bourgades » ou « villages d’abeilles »), ces apiformes sont solitaires : chaque femelle assure la construction de son propre nid et engrange l’alimentation de sa propre descendance. De nombreuses espèces d’Andrénidés sont actives dès mars et avril, collectant pollen et nectar sur les premières fleurs du printemps. Suivant les espèces, la période de vol de cette famille d’abeilles sauvages s’étend de la fin de l’hiver à la fin de l’été.
Début de l’enquête
Le fait que ce soient des andrènes femelles et non des mâles qui arboraient au front des pollinies exclut a priori la possibilité que celles-ci appartiennent aux fleurs du genre euro-méditerranéen Ophrys (qui comprend plusieurs centaines d’espèces), ces dernières n’attirant que des abeilles mâles qu’elles escroquent sexuellement (sexual deception) en imitant – via l’émission de pseudo-phéromones similaires aux phéromones sexuelles émises par les abeilles femelles de leurs espèces respectives – des abeilles femelles sexuellement réceptives. Si le mimétisme dont témoignent les orchidées du genre Ophrys est avant tout olfactif, il est aussi visuel (taille, forme, couleur et motifs ornementaux du labelle) et tactile (pilosité de ce dernier) (Wilcox, 2018).
Le jardin peuplé d’espèces florales sauvages et horticoles où furent photographiées les 3 andrènes flanquées de pollinies a été aménagé sur une partie de prairie anciennement pâturée. Le reste du pâturage (ci-dessous) constitue aujourd’hui une prairie « maigre » (non enrichie en engrais organique : fumier, lisier… ou chimique) et n’est fauchée qu’une fois par an (fin juin-début juillet).
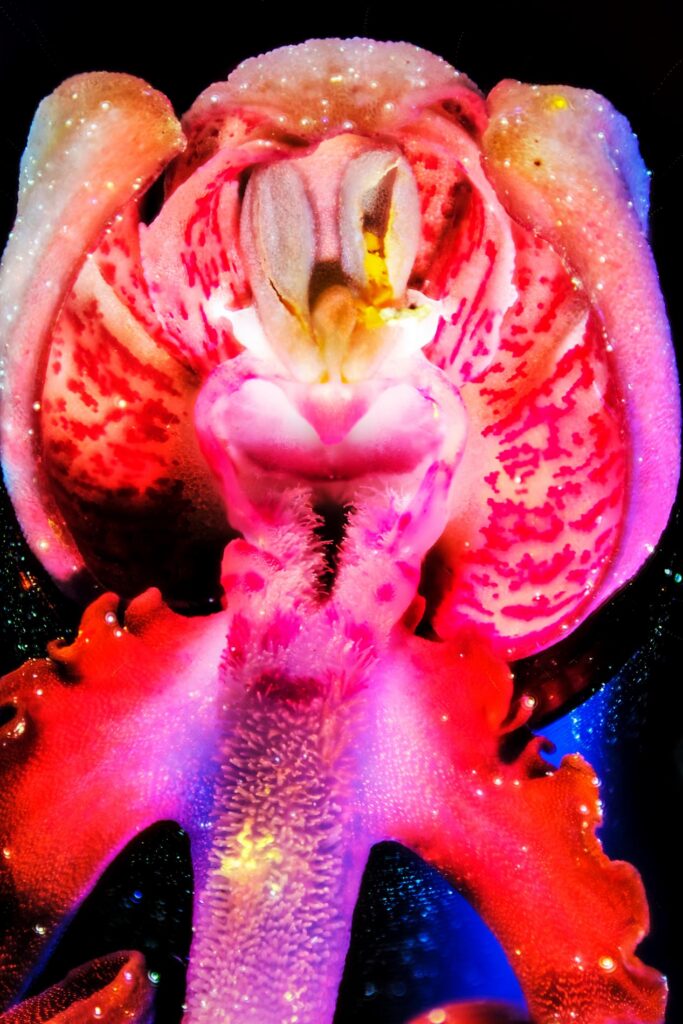
Ophrys abeille (Ophrys apifera)
En mai-juin, la flore herbacée qui y domine comprend des picris et autres Astéracées à fleurs jaunes, des marguerites et renoncules âcres (boutons d’or), du trèfle violet, diverses espèces de géraniums et de vesces, ainsi que des graminées. L’ensemble – qui compose un terrain d’un hectare – se localise dans un petit village calvadosien environné de champs intensivement gérés : de vastes parcelles dépourvues de haies, monocultivées, pesticidées et engraissées chimiquement.
Orchis pyramidal (Anacamptis pyramidalis)Orchis bouc (Himantoglossum hircinum)Une hylée, petite abeille sauvage mâle du genre Hylaeus (famille des Colletidae) se repose sur le labelle d’un Orchis bouc.
Dans le jardin comme dans la prairie naturelle (non pâturée depuis une douzaine d’années) sont disséminés quelques pieds d’Ophrys abeille (Ophrys apifera) et d’Orchis pyramidal (Anacamptis pyramidalis), tandis que nombre de pieds (plus d’une centaine) d’Orchis bouc (Himantoglossum hircinum) se concentrent dans la partie haute de l’herbage ; ces derniers poussent aussi de façon clairsemée dans le jardin. D’autres espèces d’orchidées ont été inventoriées en fleurs aux alentours du village (Orchis mascula, Dactylorhiza maculata) et dans les communes des alentours (Neottia ovata, Anacamptis morio, Epipactis helleborine, Platanthera bifolia). Pour les trois espèces déjà citées, Ophrys apifera,Anacamptis pyramidalis, Himantoglossum hircinum, elles sont aussi souvent relevées à partir de la seule présence de rosettes hivernales, les dépendances vertes villageoises, accotements routiers et bords de champs cultivés de la région subissant d’incessants broyages à l’épareuse aux mois d’avril, juin (cliché ci-dessous), août et novembre.
Carte de vœux 2020 réalisée par Yves Wilcox à partir du montage de photographies de diverses espèces d’Ophrys et de pollinisateurs avérés ou potentiels – Crédit photo : Yves Wilcox
Fond d’écran : Andrena thoracica sur Ophrys des Olonnes (proche aranifera)
De haut en bas et de gauche à droite : Diptère Myopa tessellatipennis sur Ophrys des Olonnes (proche aranifera), Andrena sp. sur O. aranifera en Saintonge, Punaise Rhinocoris erythropus et Tenthrède indéterminée sur O.aranifera en Vendée, Andrena thoracica sur Ophrys des Olonnes, Andrena carbonaria sur O. passionis en sud-Vendée, Argogorytes sp. sur O. insectifera dans la Drôme, Andrena ovatula sur O. funerea en Aveyron, Andrena sp. sur O. lupercalis en Saintonge, Andrenasp. sur O. lutea dans l’Aude, Andrena combinata sur O. aymoninii en Aveyron, Andrena sp. sur O. aranifera en Vendée.
Un expert rejoint l’enquête
Christian Noël (président de la Société française d’orchidophilie de Normandie) me conseilla d’envoyer les photos de ces 3 andrènes femelles « empollinées » à Yves Wilcox, spécialiste des orchidées et de leurs interrelations avec les insectes floricoles et membre de la Société française d’orchidophilie de Poitou-Charentes et de Vendée.
Selon Yves Wilcox, parmi les espèces observées en fleur en juin dans le secteur, aucune de ces 3 pollinies n’appartient à Ophrys apifera, à cause de son autogamie, ni à Dactylorhiza maculata, pollinisée essentiellement par des coléoptères, et des Bourdons, ni à Anacamptis pyramidalis.
Le cas d’Anacamptis pyramidalis
Base interne du labelle d’une fleur d’Orchis pyramidal – Crédit photo : Yves Wilcox
Cette dernière espèce d’orchidée est dite « trompeuse » d’un point de vue alimentaire, car elle n’exsude pas de nectar – à l’instar d’un certain nombre d’orchidées européennes ; la tromperie alimentaire (food deception) a lieu lorsqu’une plante à fleur attire des pollinisateurs sans leur prodiguer de récompense nutritionnelle. De fait, les fleurs de l’orchis pyramidal attirent des lépidoptères diurnes et nocturnes (Zygaena, Pieris, Melitaea, etc.) qui, croyant y puiser du liquide sucré, la pollinisent, leurs pièces buccales étant bien adaptées à leur morphologie : « Les plaques de guidage [de la fleur de l’Orchis pyramidal] guident la trompe [des papillons] vers l’entrée étroite de l’éperon (cliché ci-dessus). Le long éperon (ne contenant pas de nectar) et l’entrée étroite incitent le pollinisateur à enfoncer sa tête profondément dans la fleur, où il touche et repousse la bourse vers l’arrière. Le viscidium en forme de selle [plaque adhésive basale des pollinies NDLR] est une belle adaptation à la langue du papillon : lorsqu’il est touché, il s’enroule autour de la trompe, fixant fermement les pollinies qui se courbent latéralement en raison du mouvement d’enroulement du viscidium (cliché ci-dessous).En même temps, ils se penchent en avant, amenant les pollinies à la position exacte permettant de toucher les deux lobes stigmatiques à gauche et à droite de la base de la colonne. Ce processus prend environ 30 secondes, et lorsque le papillon visite une autre fleur, il poussera certainement les pollinies sur les deux lobes stigmatiques. Les papillons utilisent souvent l’inflorescence comme plate-forme d’atterrissage à partir de laquelle ils commencent à sonder les fleurs à la recherche de nectar.» (Claessens et al., 2016.)
Pollinies solidement fixées par leur viscidium sur la trompe d’un papillon – Crédit photo : Yves Wilcox
Dans l’un de ses articles, Yves Wilcox signale qu’Anacamptis pyramidalis profite de la présence de diverses plantes nectarifères prairiales tel le trèfle violet (Trifolium pratense) qu’elle imite dans sa forme et dans sa couleur afin d’escroquer les visiteurs floraux : «L’imitation du trèfle violet favorise la pollinisation par des lépidoptères. L’éperon d’A. pyramidalisne contient pas de nectar libre mais un liquide sucré non disponible pour les papillons qui est pourtant accessible à d’autres insectes après perforation. Ce liquide est probablement la trace d’une évolution vers la tromperie favorisant l’allogamie et sa production pourrait être maintenue dans l’éventualité d’un retour à la récompense. La fécondation évolue selon les milieux, les conditions climatiques et la présence des pollinisateurs. Une trop forte sélection des pollinisateurs et la tromperie peuvent conduire vers la production de nectar ou à l’autogamie, qui est un frein évolutif. L’allogamie avec imitation d’une fleur nourricière est une stratégie moins coûteuse que la production de nectar qui conserve l’avantage du brassage génétique.» (Wilcox, 2019, p.183).
Le cas d’Himantoglossum hircinum
Selon Yves Wilcox, les pollinies véhiculées par les 3 andrènes femelles photographiées sur fleurs de rue officinale et sur inflorescence de lavande appartiennent très vraisemblablement à Himantoglossum hircinum, orchidée qui pratique également le leurre alimentaire car non sécrétrice de nectar.
Crédit photo : Yves Wilcox
« Chez trois espèces d’Himantoglossum examinées, Schill & Pfeiffer (1977) ont signalé des sculptures en forme de hamule 2verruqueux. Dans le cas d’H. hircinum, on observe des portions frontales de la même massula, nettement en forme de hamule verruqueux sur les côtés, mais se fondant en gemmes de différentes tailles sur la face avant. » (Carey et al., 2002.)
Selon ce spécialiste des interactions de pollinisation entre orchidées et insectes anthophiles, la couleur orange des pollinies – dont ne subsistent que les caudicules âgées débarrassées du pollen (massulae) –collées sur la face de l’une des andrènes femelles exprime la sénescence, «fleurs et feuilles virant au jaune et au rouge par manque de chlorophylle en séchant».
Fluorescence de la fleur d’H. hircinum révélée grâce à une photographie de type UVIVF ou « fluorescence visible induite par les ultraviolets » – Crédit photo : Yves Wilcox Fleur d’Himantoglossum hircinum et pollinies – Crédit photo : Yves Wilcox
Compléments d’enquête
En guise de conclusion de ces investigations et interrogations entomo-botaniques, quelques informations collectées ci-après dans la littérature scientifique confirment les interactions de pollinisation nouées entre andrènes et Orchis bouc (dont le nom de genre Himantoglossum vient du grec himas, « lanières » et –glossa, « langue » ; le nom d’espèce hirsinum procédant du latin hircinus, « de bouc »).
Des pollinisateurs dupés… pour la bonne cause
«Dans la littérature, on mentionne une grande variété de pollinisateurs, comme les syrphes, diverses abeilles, guêpes, fourmis, coléoptères, papillons et mites. Cependant, nos observations complétées par d’autres données montrent clairement que les abeilles, surtout celles du genre Andrena, sont les pollinisateurs légitimes de H. hircinum. […] Les orchidées trompeuses sont reconnaissables à leur production irrégulière de graines : une abeille visite certaines fleurs, puis découvre qu’elle a été trompée et s’en va. Souvent, la nouaison de la moitié inférieure est plus élevée que celle de la moitié supérieure, ce qui illustre le processus d’apprentissage des abeilles inexpérimentées. La tromperie est une stratégie répandue chez les orchidées. La production de nectar nécessite beaucoup d’énergie, qui, chez les espèces trompeuses, peut être investie dans la formation des fruits. Un autre avantage est l’amélioration de la ‘‘forme physique’’ des plantes.Les orchidées à nectar sont visitées plus longtemps et les insectes ont tendance à se déplacer plus longtemps d’une fleur à l’autre sur le même épi, ce qui entraîne un degré de géitonogamie3 beaucoup plus élevé. Chez les orchidées trompeuses, les insectes visitent peu de fleurs et volent beaucoup plus vite vers la plante suivante, favorisant ainsi la pollinisation croisée, ce qui entraîne un échange de gènes plus important. Ceci est important pour maintenir une population saine.» (Claessens et al., 2016.)
«Parmi les facteurs qui auraient favorisé l’évolution de la tromperie (absence de récompense) chez les orchidées, on trouve, lorsque les pollinisateurs non récompensés visitent moins de fleurs par floraison, la réduction de la sélection par ces insectes. Nous avons obtenu des données sur les niveaux naturels de géitonogamie chez Himantoglossum hircinum en surveillant la dispersion et la réception de pollinies à code couleur. Nous avons marqué 956 fleurs de H. hircinum. 36 % des fleurs visitées par les pollinisateurs et 71 % des floraisons marquées ont connu des événements de pollinisation géitonogamique. La distance de transport du pollen la plus longue chez H. hircinum pollinisé par Andrena était de 6,9 m (médiane 1,27 m). Une analyse des études de suivi du pollen chez les orchidées a révélé des niveaux de géitonogamie d’environ 40 % chez les espèces sans récompense et les espèces récompensantes. Ces niveaux sont similaires aux niveaux de géitonogamie observés chez d’autres angiospermes pollinisés par des animaux […].
Jusqu’à présent, cependant, il n’est pas évident que les orchidées sans récompense connaissent des niveaux de géitonogamie particulièrement bas.» (Kropf et al., 2008)
Une andrène femelle (Andrena sp.) visite une fleur d’H. hircinum – Crédit photo : Yves WilcoxAbeille mellifère (Apis mellifera, Apidae) capturée par une araignée-crabe (Misumena vatia) sur une inflorescence d’H. hircinum. On notera l’homochromie de l’araignée et de l’orchidée – Crédit photo : Yves Wilcox
Orchis bouc en baie du Mont Saint-Michel- 4 juin 2016 – Dessin au pastel deMargarita Noël
«L’odorat et le goût, les plus anciens de nos sens, déterminent les composés organiques volatils émis par une source naturelle. Ils se sont probablement développés dans des organismes très primitifs comme moyen d’obtenir des informations sur les changements chimiques survenant dans leur environnement. Les animaux utilisent l’odorat et le goût pour trouver de la nourriture et évaluer sa qualité. L’odeur de la nourriture a un effet puissant sur les animaux. Les organismes vivants utilisent le sens chimique comme moyen de communication. Si la communication se fait entre différentes parties du même organisme, le messager est appelé hormone. Les produits chimiques utilisés pour transmettre des signaux d’un organisme à un autre sont connus sous le nom de produits sémiochimiques. Dans le cas des fleurs, les composants aromatiques sont principalement consacrés à attirer les insectes pollinisateurs. Ces dernières années, Himantoglossum s.l. a inclus d’autres taxons d’un intérêt et d’une conservation considérables. Actuellement, le genre élargi Himantoglossum est composé du sous-genre Himantoglossum comprenant toutes les espèces de l’ancien genre Himantoglossum, du sous-genre Barlia, composé des deux espèces du genre Barlia, et du sous-genre Comperia composé uniquement de l’ancienne espèce Comperia comperiana.
L’attraction du pollinisateur chez les orchidées se fait généralement d’abord par la diffusion aérienne des odeurs, puis par la vue, lorsque le pollinisateur s’approche de son inflorescence cible, et enfin par des signaux tactiles et des composés peu volatils et extractibles lorsqu’il se pose sur la fleur choisie. H. hircinum et H. adriaticum ont de grandes fleurs voyantes avec une longue lèvre captivante, souvent ornée de touffes voyantes de papilles colorées qui fournissent des points d’appui servant de guide aux pollinisateurs. Vöth (1990) spécule que ces papilles pourraient également être le siège d’osmophores [glandes] responsables de la plupart des composés organiques volatils émis par la plante. L’odeur émise par ces deux espèces peut être forte, désagréable ou sucrée. Ce sont des espèces allogames qui n’offrent pas de récompense alimentaire ; en fait, leur éperon court et en forme de sac ne contient pas de nectar.
Teschner (1980) a montré que l’éperon de H. hircinum et de H. adriaticum peut contenir de petites quantités de glucose chez certaines populations. Kropf et Renner (2008) ont démontré chimiquement la présence de nectar dans H. hircinum. On sait peu de choses sur la pollinisation des différentes espèces du groupe H. hircinum ; on pense que les pollinisateurs diffèrent localement, Teschner (1980) suggérant que les abeilles solitaires en sont les véritables pollinisateurs. […] Le parfum d’une fleur peut être un facteur important déterminant la pollinisation d’une plante. L’étude du parfum des orchidées a fait l’objet de plusieurs travaux dans le passé. Malheureusement, plusieurs approches différentes ont été utilisées afin de déterminer la composition de l’arôme d’une orchidée. […] Souvent, différentes procédures chimiques ont permis d’obtenir des résultats différents. (Mecca et al., 2021.)
En 2025…
Deux autres espèces d’andrènes furent photographiées à la mi-juin au sein du même jardin, flanquées de pollinies d’Orchis bouc tout proches alors en fleur.
Ci-dessous : Une andrène mâle (Andrena sp.) porteuse d’une paire de pollinies d’H. hircinum est retrouvée morte dans une rose… un beau cercueil quand même.
Ci-dessus : Deux andrènes de la bryone femelles (Andrena florea) arborent au front une paire de pollinies d’H. hircinum pour l’une, deux paires pour l’autre.Cinq grêlons de la taille d’une noix parmi ceux tombés en rafale après mi-juin 2025 sur la localité – Crédit photo : Florence Thérèse
Malheureusement, quelques jours à peine après que ces photos furent prises, un orage de grêlons énormes (jusqu’à 5 cm) tombés en rafales décima mon troupeau d’Orchis boucs et quantité d’autres herbacées en pleine floraison ou sur le point de l’être. Quant aux arbres, la plupart d’entre eux perdirent un tiers à la moitié de leurs feuilles dans la tourmente… et bien sûr leurs fruits naissants, pour les arbres fruitiers… Du jamais vu dans cette région de l’ex-Basse-Normandie à la mi-juin… Avant que n’arrivent les fortes chaleurs qui firent de ce mois de juin 2025 le 2e mois de juin le plus chaud jamais enregistré avec une anomalie de +3,3 °C, derrière juin 2003 (+3,6 °C)… Sale temps pour les plantes et leurs pollinisateurs…
Teschner, W. 1980. Sippendifferenzierung und Bestäubung bei Himantoglossum Koch, in : Senghas, K. & Sundermann, H., eds. Probleme der Evolution bei europaïschen und mediterranen Orchideen, pp. 104–115. Orchidee Sonderheft (Special Issue).
Wilcox Y., 2018. Pollination in the Ophrys (L.) genus : cephalic and abdominal pseudocopulations. L’Orchidophile 216: 69-80.
Wilcox Y., 2019. Pollination of Anacamptis pyramidalis : morphological adaptations and attraction strategies. L’Orchidophile 221: 171-184.
Wikipedia consacre un excellent article aux « fleurs cadavres » ou fleurs sapromyophiles (du grec sapros, « putride », myia, « mouche » et philia, « ami de »), plantes « dont la fleur émet des odeurs stimulant l’attraction d’insectes pollinisateurs saprophages, coprophages et nécrophages (diptères, coléoptères) exercée par des chairs en putréfaction, du fumier, de la matière fécale, de l’urine ou des champignons, typiques des milieux de ponte où ces animaux ont l’habitude de déposer leurs œufs donnant des larves qui s’en nourrissent. »
Cet article est adapté d’une première parution début 2025 dans les pages du Bulletin de la Société française d’orchidophilie de Normandie. Avec l’aimable autorisation de sa rédaction.
L’autrice remercie pour leur collaboration Christian Noël, Michel Beer, Margarita Noël, Jean-François Odoux et surtout Yves Wilcox, dont l’expertise orchidophile a permis l’aboutissement de cet article.
En entomologie, ce terme signifie « pièce en forme de crochet des ailes des insectes hyménoptères ». En botanique, « hamuleux » a pour sens : « Qui est pourvu de petits poils crochus. » ↩︎
La pollinisation est de deux types : l’autopollinisation (autogamie) et la pollinisation croisée (xénogamie, allogamie). L’autogamie consiste dans le dépôt de grains de pollen sur le stigmate de la même fleur (fleur hermaphrodite), tandis que la géitonogamie consiste dans le dépôt de grains de pollen sur le stigmate d’une autre fleur de la même plante (pollinisation de voisinage). Le dépôt de grains de pollen sur le stigmate d’une fleur d’une autre plante est appelé xénogamie. L’autogamie et la géitonogamie sont deux types d’autopollinisation dans l’étude de la génétique. Mais fonctionnellement, la géitonogamie est un type de pollinisation croisée. La géitonogamie se différencie ainsi des modes de reproduction que sont xénogamie et autogamie. D’un point de vue génétique, si la fécondation géitonogame réussit, elle a le même effet que l’autopollinisation, car il n’y a pas de distribution ou de recombinaison du matériel génétique (idiogamie, consanguinité, au contraire d’une pollinisation croisée). Du point de vue de la variabilité génétique, la géitonogamie équivaut donc à l’autogamie.Si une plante est auto-incompatible (comme l’est le noyer d’Amazonie), la géitonogamie réduit la production de graines. ↩︎
Adepte du Spipoll, Isabelle Avisse écrit pour diverses revues : La Santé de l’abeille, L’Orchidophile, le Bulletin de la Société française d’orchidophilie de Normandie, Abeilles en liberté. Ancienne apicultrice, auteure d’une exposition sur les abeilles sauvages, elle l’est également d’un Grand traité des miels publié en 2014 aux éditions Le Sureau. Contact :